2.4 Antibody activity of IgG subclasses
The IgG subclass distribution in specific antibody responses has been found to vary with structure of the antigen (nature of carrier, number and nature of the epitopes, physicochemical properties), its dose and route of entry, as well as with genetic constitution of the host. In contrast to T cell-independent (thymus-independent) antigens, T cell-dependent thymus-dependent) antigens require interaction with helper T lymphocytes in order to stimulate B-lymphocytes to antibody production. Interestingly, stimulation of antibody responses towards certain antigens may result in a selective increase in IgG antibodies of certain subclasses (34,35,36). Whereas antibodies against bacterial and viral protein antigens such as tetanus toxoid or outer-membrane components, which are T cell-dependent antigens, can be detected in all four IgG subclasses, IgG1 is the prevailing isotype, sometimes in combination with IgG3(37). Anti-protein antibodies of the IgG2 subclass generally provide only a marginal contribution. On the other hand, IgG antibodies against polysaccharide antigens, which are generally T cell-independent, generally show a much more pronounced subclass distribution: immunization with several encapsulated bacteria leads to an almost exclusive IgG2 anti-polysaccharide response (38). An exception is seen in children under the age of 2-3 years, in whom anti-polysaccharide antibodies have been found to occur in the IgG1 subclass (39).
Repeated, long-term antigenic stimulation with T cell-dependent antigens may lead to a marked IgG4 antibody response(40). In general, anti-viral IgG antibodies are highly restricted to IgG1 and IgG3, with IgG3 antibodies appearing first in the course of infection. The IgG subclass distribution in an anti-bacterial response will be more heterogeneous, since bacteria contain many different antigenic epitopes, with considerable variations in their protein and carbohydrate structures.
The IgG subclass determination of antigen-specific antibodies is still quite cumbersome, although many studies are performed in this area. As a major drawback, no generally accepted international age-related reference values are available. Moreover, different assay systems do not always provide consistent results. The antigen-specific antibodies are mostly determined by means of ELISA, the antigen being coated on a microtiter plate, followed by incubation with the antibodies to be characterized, and finally with enzyme-labelled monoclonal anti-IgG subclass-specific antibody.
2.5 Effector functions of IgG subclasses
The most important biological activities of antibodies are related to their effector functions, aimed at inactivation or removal of infectious agents and their products (e.g. bacteria, viruses, toxins). Antibodies of the IgG class exert two major effector functions: activation of complement and opsonisation (i.e. the induction of phagocytosis). These effector functions, mediated via the (constant) Fc fragment are induced as a result of interaction of the antibody with its antigen via the (variable) Fab moiety.
The four IgG subclasses differ from each other with respect to their effector functions (table II). This difference is related to differences in structure, notably with respect to the interaction between the variable, antigen-binding Fab-fragments and the constant Fc fragment (figure 4). In particular, the length and flexibility of the hinge region are different. The flexibility of the hinge region decreases in the order IgG3>IgG1>IgG4>IgG2. This probably relates to the higher activity of IgG3 in triggering effector functions, when compared to the other subclasses (21,41).
2.5.1 Complement activation
Serum bactericidal activity is induced when the complete complement cascade (C1 to C9) is activated. The formation of the membrane attack complex (MAC) is the final result of activation by the classical pathway or the alternative pathway of complement. Unlike the classical pathway, activation of the alternative pathway is not dependent upon antibodies, although many indications exist for an enhancing activity of antibodies in the alternative pathway. The MAC (C5b-9) is inserted into the membrane of target cells, after which these are killed by a lytic mechanism. Besides, opsonisation of micro-organisms by complement components (C3b, iC3b and C3dg) may result in phagocytosis. The structural diversity in the hinge region of IgG subclasses entails differences in their ability to activate complement. The initial event in the activation of the complement cascade by the classical pathway is the binding of C1q to sites present on the CH2 domains of IgG. This binding occurs when the latter is aggregated, usually through formation of an antigen-antibody complex. Other components required for complement activation include a disulfide bond in the amino-terminal end of CH2 (normally the hinge region), the presence of carbohydrates and paired C(H)3 domains. The capacity of the four human IgG subclasses (in monomeric form) to bind C1q decreases in the order: IgG3>IgG1>IgG2>IgG4 (5,31). IgG4 does not activate complement. Hinge-dependent Fab-Fab and Fab-Fc flexibility determines the accessibility of the complement binding site to C1q.IgG3, the human IgG subclass with the longest hinge, is the most effective complement activator. The inability of IgG4 to activate complement is caused by the structure of the Fc fragment as well as the steric hindrance of the complement binding sites by the Fab arms (figure 5).
Although IgG2 is a poor activator of the classical complement cascade, effective opsonisation and killing of certain bacterial strains and protection in bioassays by purified IgG2 have been reported. This may be explained by a combined effect of both bacterial antigen and IgG2 on the activation of complement. Density and accessibility of antigenic determinants on the bacteria may determine the actual complement activation by IgG subclasses. Alternatively, IgG2 may utilize the alternative pathway in complement activation in a Fc-independent way, bypassing C1q binding (table II) (42,43,44). Complement activation via the alternative route appears to be especially important in the opsonisation and killing of bacteria.
2.5.2 Opsonisation and induction of phagocytosis
Most micro-organism are by themselves incapable to activate complement or to bind phagocytes. Antibodies act as flexible adapters mediating the adherence of infectious agents to phagocytes. The antibody forms a bridge to attach the microbe to the Fc receptor on the phagocyte membrane.
Interaction with Fc receptors
Phagocytosis is initiated by an interaction between the Fc fragment of the immunoglobulin and receptors. Fc receptors for IgG (FcgR) are expressed primarily on effector cells of the immune system, in particular macrophages, monocytes, myeloid cells and dendritic cells.
The four human IgG subclasses show differences in their interaction with FcgR's. Binding of the Fc part of IgG to a FcgR is instrumental in the induction of the cell's effector function (45, 46). In this way, FcgR's play a key role in bridging (IgG) antibody activity and cellular effector mechanisms. The latter comprise e.g. phagocytosis, endocytosis, antibody-mediated cellular cytotoxicity, release of a range of inflammatory mediators, antigen presentation and clearance of immune complexes. Moreover, since several FcgR-bearing cell types (e.g. macrophages, monocytes and dendritic cells), have the capacity to present antigens to T lymphocytes, FcgR-induced phagocytosis also plays a role in antigen presentation and amplification of the immune response. As a result, interaction between FcgR and IgG antibodies is pivotal in the immune response against infectious agents (47). Since IgG subclasses differ in their affinity to FcgR, clinical manifestations of IgG subclass deficiencies will depend upon the properties of the FcgR as well.
Different types of human FcgR
It has been shown that on human leukocytes three distinct types of Fcg -receptors occur, which are distinguishable by structural and functional properties, as well as by antigenic structures detected by CD monoclonal antibodies. They are designated FcgRI, FcgRII, and FcgRIII, respectively and are differentially expressed on (overlapping) subsets of leukocytes (table II) (48).
FcgRI (CD64), a high-affinity receptor expressed on monocytes, macrophages, neutrophils, myeloid precursors and dendritic cells, comprised isoforms la and lb. Fcg RI has a high affinity for monomeric human IgG1 and IgG3. Its affinity for IgG4 is about 10 times lower, while it does not bind IgG2. Fcg RI does not show genetic polymorphism (48).
FcgRII (CD32), comprised of isoforms IIa, IIb1, IIb2, IIb3 and IIc, is the most widely distributed human Fcg R type, being expressed on most types of blood leukocytes, as well as on Langerhans cells, dendritic cells and platelets. FcgRII is a low-affinity receptor, that only binds aggregated IgG. It is the only Fcg R class able to bind IgG2. FcgRIIa shows genetics polymorphism, resulting in two distinct allotypes, Fcg RIIa-H131 and Fcg RIIa-R131, respectively (49,50). This functional polymorphism is attributable to a single amino acid difference: a histidine (H) or an arginine (R) residue at position 131, which is critical for IgG binding. FcgRIIa readily binds human IgG and IgG3 and appears not to bind IgG4. The Fcg RIIa-H131 has a much higher affinity for complexed IgG2 than the Fcg RIIa-R131 allotype (50).
Fcg RIII (CD16) has two isoforms, both of which are able to bind IgG1 and IgG3. The FcgRIIa, with an intermediate affinity for IgG, is expressed on macrophages, monocytes, natural killer (NK) cells and subsets of T cells. FcgRIIIb is a low-affinity receptor for IgG, selectively expressed on neutrophils. It is a highly mobile receptor with efficient collaboration with other membrane receptors. Studies with myeloma IgG dimers have shown that only IgG1 and IgG3 bind to FcgRIIIb (with low affinity), while no binding of IgG2 and IgG4 has been found (51,52). The FcgRIIIb bears a co-dominant, bi-allelic polymorphism, the allotypes being designated NA1 (Neutrophil Antigen) and NA2(53).
FcgR genetic polymorphism, IgG subclasses and disease In human, IgG2 antibodies have been found to play a key role in immunity against infection with encapsulated bacteria. This defense does not only depend upon complement activation, since IgG2 has a poor complement-activating activity, at least when the epitope density is low. Indeed, it has been shown that phagocytosis by neutrophil granulocytes is the major underlying mechanism. Moreover, the FcgRIIa polymorphism is important, neutrophils from FcgRIIa-H131 homozygous individuals phagocytising better than those of FcgRIIa-R131 individuals (54). This polymorphism will be more critical when IgG2 levels are low, as is the case in children. In children surviving meningococcal septic shock, the allotype FcgRIIa-R131 occurred twice as often as in normal donors (47).
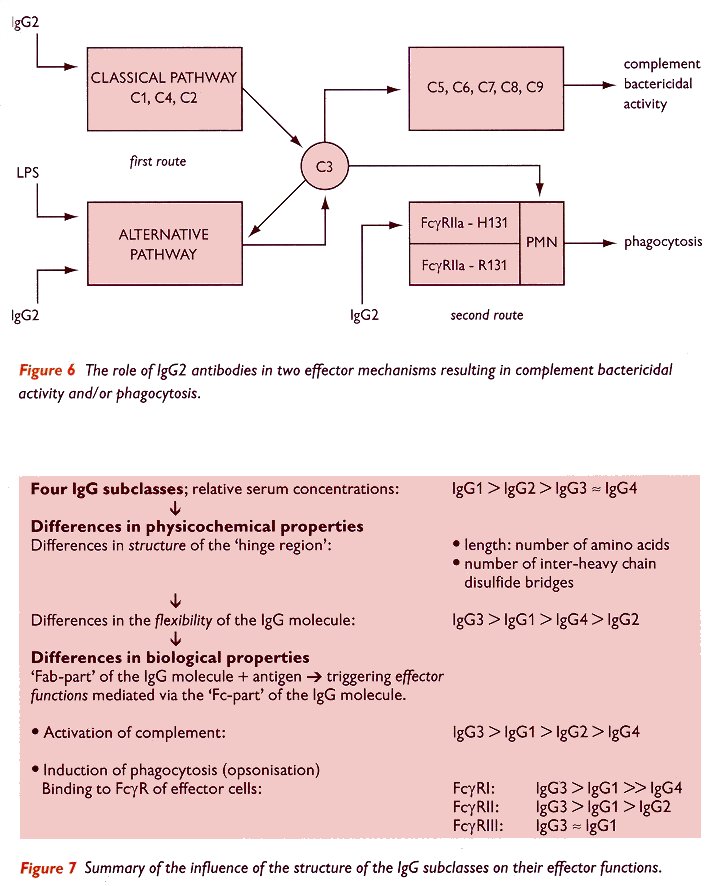
Figure 6 (bottom of page) schematically illustrates the role of IgG2 in the defense against micro-organisms and the different underlying routes The first route involves the activation of complement, either by the classical pathway (C1, C4 and C2), or by the alternative pathway (which is potentiated by antibodies of the IgG2 subclass, whereas it should be kept in mind that IgG2 is a poor activator of the classical pathway). In the alternative pathway, bacterial components (indicated here as lipopolysaccharide, LPS) also play an essential role. Activation of the complement cascade, either by the classical or the alternative pathway, will lead to the activation of complement component C3. The binding of activated C3 to the bacterial membrane will lead either to direct killing (mediated by the complement cascade C5 to C9), or to opsonisation resulting in binding to complement receptors on polymorphonuclear cells (PMN), followed by phagocytosis.
According to the second route indicated in figure 6, bacteria sensitized by IgG2 antibodies may also be phagocytised when they bind to the FcgRIIa on the cell membrane of the PMN. This latter process will only be effective in individuals expressing a FcgRIIa of the allotype H131, since the other allele, Fcg RIIa-R131, is unable to bind IgG2 effectively.
With respect to FcgRIIIb polymorphism, it was reported that the combined FcgRIIa-R131 and FcgRIIIb-NA2 phenotype is associated with meningococcal infection (55).
Thus, next to a quantitative defect of IgG subclass levels, effector functions of humoral immunity may also be negatively affected by defects in Fc receptors